Introduction
Sepsis is a type of systemic inflammatory syndrome caused by infections. It commonly leads to an unbalanced host response to infection, inducing organ dysfunction [1]. Sepsis remains one of the leading causes of mortality worldwide. The global annual incidence of sepsis was found to be 276-678/100,000 persons. Case fatality ranged from 22.5% to 26.7% [2]. In addition, one-third of patients admitted to intensive care units survive for < 30 days, and mortality rates increase with age, disease status, and the type of organ dysfunction. Patients who survive sepsis have longer rehospitalization times and a higher risk of subsequent mortality. Patients with early sepsis often have definitely suspected infections. If not detected and treated early, it is likely to progress to septic shock, leading to adverse outcomes, such as mortality [3, 4]. Thus, seeking early diagnostic indicators and novel therapeutic targets for sepsis has become a research hotspot.
Chitinase-3-like protein 1 (CHI3L1) is a secreted glycoprotein. It belongs to the glycoside hydrolase family 18. It is located on chromosomes 1q31-q32, at the same site as receptor genes for C-reactive protein and interleukin (IL)-6 [5]. CHI3L1 has chitin binding activity. Chitin is a key compound that promotes cell proliferation in the body. Therefore, CHI3L1 is also considered to have cytokine and growth factor properties [6]. CHI3L1 is synthesized and secreted by a variety of cells, including macrophages, neutrophils, synovial cells, chondrocytes, fibroblast-like cells, smooth muscle cells, and tumor cells [7]. It plays a crucial role in tissue damage, inflammation, tissue repair, and remodeling responses. CHI3L1 has been linked to the development of Alzheimer’s disease, osteoporosis, and cancer [8-10]. Its abnormal expression usually affects the action of inflammatory cytokines (IL-1β, IL-6, and tumor necrosis factor α [TNF-α]). Recent studies have elucidated the role of CHI3L1 in sepsis, demonstrating its elevated expression during systemic inflammation and its association with disease severity. Elevated levels of CHI3L1 have been shown to be associated with poor outcomes in septic patients, indicating its potential as a prognostic biomarker [11, 12]. However, the specific regulatory mechanisms and the complete molecular mechanisms of action of CHI3L1 are not yet fully understood.
Lactate (LA) is an intermediate product of glycolytic metabolism. A clinical trial has demonstrated that LA levels are associated with organ failure in patients with sepsis [13]. Lactylation is a post-translational modification (PTM). Specifically, LA accumulated during metabolism in the body functions as a precursor substance to lactylation protein lysine. This PTM regulates protein expression by altering its properties [14]. Thus, lactylation is widely involved in a variety of life activities. Lactylation has been shown to be involved in the progression of myocardial infarction, tumors, pulmonary fibrosis, and other diseases [15-17]. Studies have demonstrated that lactylation of proteins can affect the progression of sepsis [18-20]. However, there is currently no literature reporting the relationship between CHI3L1 and lactylation modification, which represents a key innovation of this study.
Collectively, the present study aimed to explore the specific mechanisms and regulatory approaches of CHI3L1 in sepsis in vivo and in vitro. It was hypothesized that CHI3L1 is modified by lactylation to promote sepsis.
Material and methods
Clinical sample collection
Blood samples were collected from 50 patients (18-60 years old) with sepsis and 50 healthy volunteers (18-60 years old) between November 1, 2023 and June 30, 2024. All subjects were diagnosed with sepsis at Sinopharm Tongmei General Hospital. The diagnosis was based on a sequential organ failure assessment score ≥ 2 [21]. All participants signed informed consent forms prior to sample collection. The present study was approved by the Ethics Committee of Sinopharm Tongmei General Hospital (No. 2023-K-07). Clinical blood samples were handled in strict accordance with the ethical standards of the Declaration of Helsinki.
Blood sample collection
Peripheral venous blood samples were collected using an EDTAK collection vessel (Baokang, Liaoning, China). Whole blood was stored at 4°C. Following centrifugation at 3,000 r/min for 10 min, the serum samples were placed in a 1.5 ml enzyme-free centrifuge tube (Corning, Inc.). The serum samples were stored at –80°C for use in follow-up experiments.
Cell culture and transfection
The mouse monocyte macrophage cell line (RAW-264.7) was purchased from the Cell Resource Center (Beijing, China). RAW264.7 cells were cultured in DMEM containing 10% fetal bovine serum and 1% penicillin. The cells were placed in a constant temperature and humidity incubator at 37°C with 5% CO2. When the cell confluency reached 80%, the cell density was adjusted to 5 × 104/ml. The cells were then inoculated into 24-well plates. After the cell growth and fusion reached 50%, 10 nM small interfering RNA CHI3L1 (si-CHI3L1#1 and #2) or its negative control (si-NC) was transfected into the RAW264.7 cells using Lipofectamine 3000 reagent (Thermo Fisher Scientific, Inc.) according to the manufacturer’s instructions. After 24 h, the transfection efficiency was detected using reverse transcription-quantitative PCR (RT-qPCR).
si-CHI3L1#1: sense CAACCUUUCCUGUGCACCCACAUCA; antisense UGAUGUGGGUGCACAGGAAAGGUUG;
si-CHI3L1#2: sense GAGUCGAACUAUGACAAGCUGAAUA; antisense UAUUCAGCUUGUCAUAGUUCGACUC;
si-NC: sense CAAUUCUUCUGCGCACCCA- ACCUCA; antisense UGAGGUUGGGUGCGCAG- AAGAAUUG.
Cell grouping and treatment
The RAW264.7 cells were inoculated in six-well plates and grew to 60-80% confluency. The cells underwent serum starvation for 12 h. These cells were divided into four groups as follows: the control group, lipopolysaccharide (LPS) group, LPS + LA group, and LPS + 2-deoxy-D-glucose (2-DG, a lactate inhibitor [22]) group. To establish the cell model of sepsis-induced injury, 1 µg/ml LPS was added to the cell culture medium and incubated with the cells for 24 h. In addition, 10 µM LA (MilliporeSigma) and 2-DG (MilliporeSigma) were incubated with the cells for 24 h. The cells treated with the same amount of PBS for 24 h were used as the control group.
Determination of LA content
The LA content in the cells was determined using a Lactate Assay Kit (MilliporeSigma). All operations were strictly carried out in accordance with the instructions provided with the kit.
RT-qPCR
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used to extract total RNA from the serum samples and cell lines. The BeyoRT III M-MLV kit (Beyotime Institute of Biotechnology) was used for RNA reverse transcription. The One-Step SYBR PrimeScript RT-PCR kit (Takara Biotechnology Co., Ltd.) was used to perform qPCR analysis on the CFX96 TOUCH real-time PCR instrument (Bio-Rad Laboratories, Inc.). The 2-ΔΔCq method was used to calculate the mRNA expression. GAPDH was used as the internal standard. The primers used in the present study were as follows: CHI3L1 (human) forward 5’-GTGAAGGCGTCTCAAACAGG-3’, reverse 5’-GAAGCGGTCAAGGGCATCT-3’; CHI3L1 (mouse) forward 5’-ATGCACACCTCTACTGAAGCC-3’, reverse 5’-ACCAGCTTGTACGCAGAGC-3’; GAPDH (human) forward 5’-GGAGCGAGATCCCTCCAAAAT-3’, reverse 5’-GGCTGTTGTCATACTTCTCATGG-3’; GAPDH (mouse) forward 5’-AGGTCGGTGTGAACGGATTTG-3’, reverse 5’-GGGGTCGTTGATGGCAACA-3’.
Western blot analysis
RIPA lysis buffer (Beyotime Institute of Biotechnology) was used to obtain total proteins from cells. A Pierce BCA Protein Assay Kit (Invitrogen; Thermo Fisher Scientific, Inc.) was used to measure protein concentration. SDS-PAGE (Thermo Fisher Scientific, Inc.) was used to isolate proteins. Following constant pressure electrophoresis, proteins were transferred to the Immobilon-E PVDF membrane (Merck KGaA). After washing the membrane with PBS solution three times, the membrane was incubated with BSA solution for 2 h at room temperature. The membrane was cultured with the primary antibodies at 4°C. Subsequently, goat anti-rabbit IgG H&L (HRP) was incubated with the membrane for 2 h at room temperature. Following repeated washing with PBST (phosphate-buffered saline with Tween), the blot was visualized using the enhanced ECL chemiluminescent substrate kit (Shanghai Yeasen Biotechnology Co., Ltd.). The antibodies used in the present study included the following: anti-CHI3L1 (1 : 2,000, Abcam), anti-p-IkB-a (1 : 1,000, Abcam), anti- IkB-a (1 : 1,500, Abcam), anti-p-p65 (1 : 1,200, Abcam), anti-p65 (1 : 1,500, Abcam), anti-Pan Lactic acid-lysine rabbit pAb (1 : 2,000, ABclonal Biotech Co., Ltd.), anti- GAPDH (1 : 3,000, Abcam) and goat anti-rabbit IgG H&L (HRP) (1 : 2,500, Abcam).
Protein degradation analysis
Protein degradation assays were based on the use of the protein synthesis inhibitor, cycloheximide (CHX). RAW264.7 cells were treated with CHX (100 µg/ml). The cells were collected at various time points (0, 6, 12, and 24 h) following treatment with CHX, and whole cell lysate was prepared and analyzed for CHI3L1 expression using western blot analysis.
Enzyme-linked immunosorbent assay (ELISA)
The IL-1b, IL-6, and TNF-a ELISA kits were purchased from Biovision. The experiment was carried out according to the manufacturer’s instructions.
Apoptosis assay
RAW264.7 cells were incubated at 37°C for 24 h. The cells were centrifuged for 2 min and the supernatant was removed. After adding 500 µl of binding buffer, 5 µl of 7-AAD solution, and 5 µl of Annexin V-PE solution (Liankebio), the cells were incubated in the dark for 30 min. Apoptosis was detected using flow cytometry.
Cell Counting Kit-8 (CCK-8) assay
A CCK-8 kit (Beyotime Institute of Biotechnology) was used for cell viability detection. According to the manufacturer’s protocol, the transfected RAW264.7 cells were incubated with 10 µl of CCK-8 solution in DMEM for 4 h. The absorbance was measured at 450 nm using a microplate reader.
Immunofluorescence (IF) staining
The cells were washed with PBS, fixed with 4% paraformaldehyde for 20 min, permeabilized with 0.5% Triton X-100 (Beyotime Institute of Biotechnology), blocked with 5% BSA, and then incubated with primary antibody (anti-CHI3L1 or anti-p65) overnight at 4°C. This was followed by incubation with goat anti-rabbit IgG H&L (Alexa Fluor 488) (cat. no. ab150079, Abcam) (green) for 2 h. Subsequently, 4-6-diamidino-2-phenylindole (DAPI, Beyotime Institute of Biotechnology) (blue) was used to stain the nuclei. Finally, images of the cells were obtained using a confocal microscope (Leica Microsystems GmbH).
Animal experiments
The animal experiments were approved by the Ethics Committee of our hospital (no. 2023-K-07). SPF healthy male C57BL/6 mice (8 weeks old) were purchased from the National Rodent Laboratory Animal Resource Bank (Shanghai, China). The mice weighed ≥ 200 and < 250 g. The mice were fed in feeding boxes at 25°C and a humidity of 50-60%. The light and dark cycle time of the feeding tank was 12 : 12 h. Mice had free access to feed and water. The present study strictly followed the Animals (Scientific Procedures) Act 1986. At the same time, all the steps involved in the present study were strictly approved by the Animal Protection and Utilization Committee of Sinopharm Tongmei General Hospital.
The mice were randomly divided into four groups (6 mice in each group) as follows: the control group, model group, model + Lv-sh-NC group, and model + Lv-sh-CHI3L1 group. Apart from the control group, the other mice were administered intrathecal injections with 10 mg/kg LPS to establish the model of sepsis. The mice in the control group were injected with the same amount of normal saline. At 1 day prior to the LPS injection, the mice were anesthetized by an intraperitoneal injection of 2% pentobarbital sodium (40 mg/kg) and then administered intrathecal injections of sh-CHI3L1 and its negative controls. All of the lentiviruses used in this experiment (Lv-sh-NC and Lv-sh-CHI3L1) were generated using the pLKO.1-puro vector and provided by OBIO Technology (Shanghai, China). After 24 h, all mice were euthanized by an intraperitoneal injection of 2% pentobarbital sodium (160 mg/kg). The heart, liver, lung, and kidney tissues and blood samples were collected. 0.5 ml blood samples were collected from each mouse through jugular vein sampling.
sh-CHI3L1 sense 5’-TCTGTTGGAGGGTGGAA- ATTT-3’, antisense 5’-AAATTTCCACCCTCCAACAGA-3’;
sh-NC sense 5’-GCAAGCTTATCGGCCAAG- CTTGAAGACGC-3’, antisense: 5’-GCGTCTTCAAGC- TTGGCCGATAAGCTTGCA-3’.
Liver and renal function evaluation
Renal function was assessed by detecting creatinine (Cr) and blood urea nitrogen (BUN) activities [23]. Liver function was evaluated by detecting alanine aminotransferase (ALT) and aspartate transaminase (AST) activities [24]. Mouse serum samples were collected to measure the serum levels of Cr, BUN, ALT, and AST using corresponding reagent kits, provided by the Nanjing Jiangcheng Bioengineering Institute.
Hematoxylin and eosin staining
The mouse tissues (heart, liver, lung, and kidney) were fixed using 4% paraformaldehyde for 24 h, paraffin-embedded, and sliced at a thickness of 4 µm. These sections were stained with hematoxylin for 5 min and eosin for 2 min. A BX51 optical microscope (Olympus Corp.) was used to observe the sections.
Statistical analyses
SPSS 20.0 software was used to perform the statistical analyses. Data are presented as mean ± standard deviation (SD). Statistical differences among groups were calculated using Student’s t-test and one-way ANOVA followed by the Tukey test. The potential diagnostic value of CHI3L1 in sepsis was assessed by receiver operating characteristic (ROC) curve analysis. A value of p < 0.05 was considered to indicate a statistically significant difference.
Results
CHI3L1 is highly expressed and LA levels are increased in sepsis
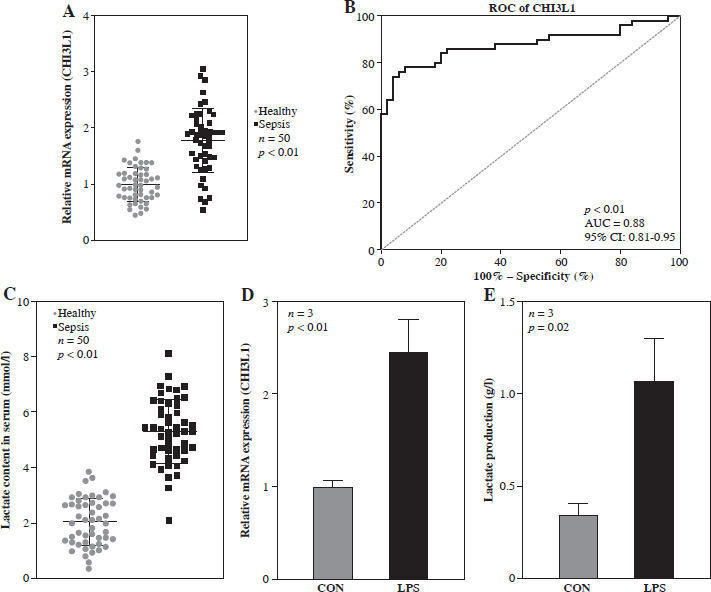
In order to explore the function of CHI3L1 in sepsis, the expression of CHI3L1 was examined in blood samples of patients with sepsis and compared with that in normal healthy participants. The results of RT-qPCR revealed that the mRNA expression of CHI3L1 was significantly elevated in the serum of patients with sepsis (Fig. 1A). The ROC analysis demonstrated that the area under the curve (AUC) of CHI3L1 was 0.88 (95% CI: 0.81-0.95), indicating that CHI3L1 may be a sensitive biomarker for sepsis prognosis and diagnosis (Fig. 1B). As shown in Figure 1C, the LA content was significantly elevated in the serum of patients with sepsis. Subsequently, a cell model of sepsis was established using LPS to treat RAW264.7 cells. The results of RT-qPCR revealed that the mRNA expression of CHI3L1 and LA production were significantly elevated in LPS-exposed RAW264.7 cells (Fig. 1D, E). These data indicated that CHI3L1 expression and LA production level were elevated in patients with sepsis and in the cell model of sepsis.
Fig. 1
High levels of CHI3L1 were associated with sepsis. A) mRNA expression of CHI3L1 in blood samples (p < 0.01). B) ROC analysis used to evaluate the diagnostic value of CHI3L1 expression in sepsis. C) Lactate concentration in sepsis patients (p < 0.01). Expression level of CHI3L1 (D) and LA content (E) in RAW264.7 cells treated with LPS
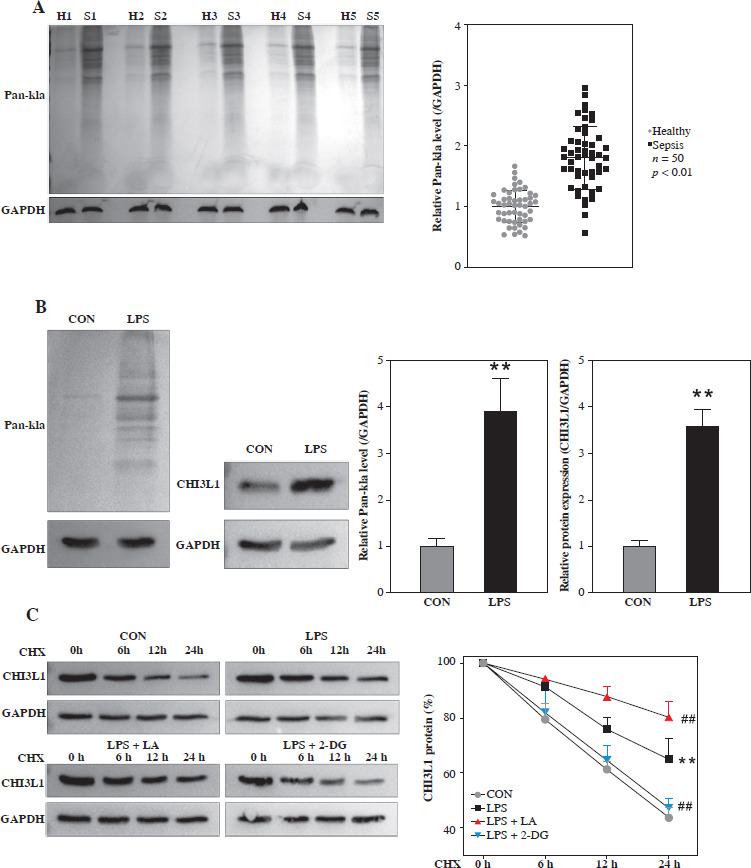
Lactation regulates the protein stability of CHI3L1
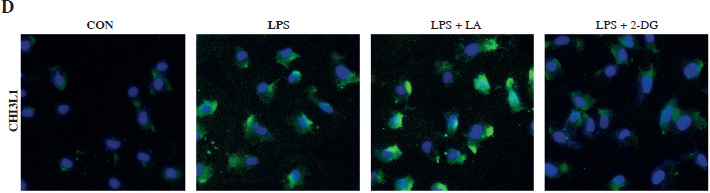
The study then analyzed the lactylation of CHI3L1 in the cell model of sepsis. As shown in Figure 2A, the level of lactylation modification in blood samples of sepsis patients was elevated. In the cell model of sepsis, the overall lactylation modification and the CHI3L1 lactylation level were also significantly elevated (Fig. 2B). Subsequently, the LPS-exposed Raw264.7 cells were treated with the protein synthesis inhibitor CHX to assess protein stability. As illustrated in Figure 2C, the degradation rate of CHI3L1 protein in the LPS group was lower than that in the control group. Treatment with LA further reduced the degradation rate of CHI3L1, whereas 2-DG promoted the degradation of CHI3L1. The results of IF staining revealed that the CHI3L1 protein levels increased following exposure to LPS. LA further increased the CHI3L1 levels, whereas 2-DG reduced the CHI3L1 levels in LPS-exposed RAW264.7 cells (Fig. 2D). These results suggested that CHI3L1 underwent lactylation in LPS-exposed Raw264.7 cells, and the stability of CHI3L1 was enhanced.
Fig. 2
Lactate regulated the stability of CHI3L1 protein. A) Lactylation level in blood samples of sepsis patients detected by western blot. B) Overall lactylation level of RAW264.7 cells treated with LPS and protein level of CHI3L1 after LPS treatment detected by western blot. C) Stability changes of CHI3L1 proteins in LPS stimulated RAW264.7 cells treated with LA and glycolysis inhibitors (2-DG) for 24 h. **p < 0.01, ##p < 0.01 D) CHI3L1 expression in LPS stimulated RAW264.7 cells treated with LA and glycolysis inhibitors detected by IF.
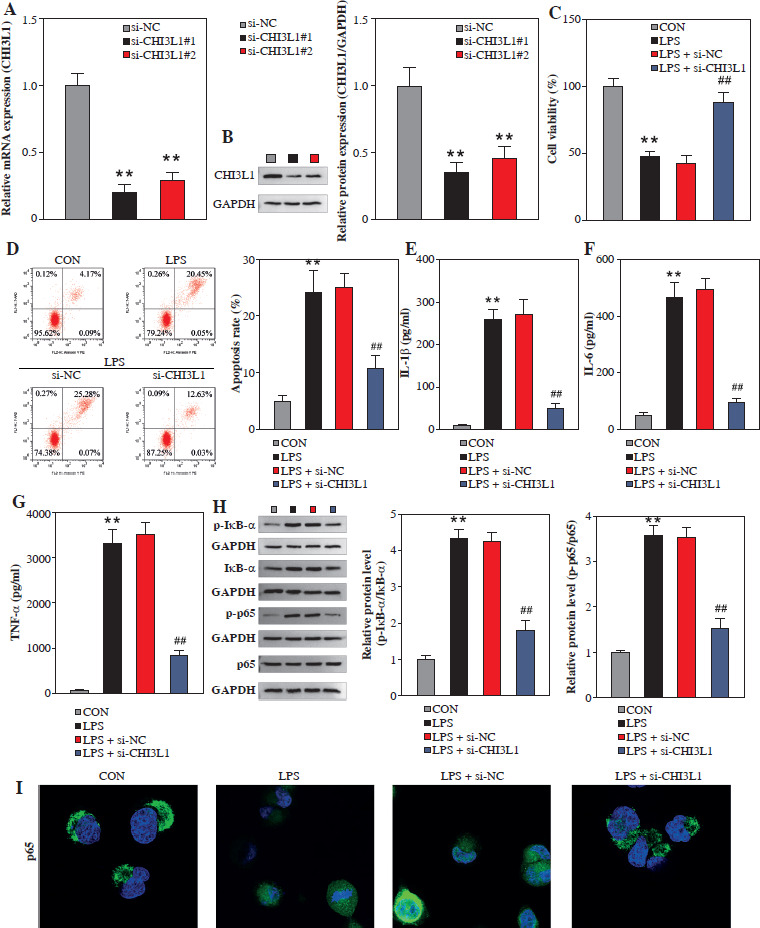
Knockdown of CHI3L1 inhibits apoptosis of cells in the cell model of sepsis
To investigate the effects of CHI3L1 on cell biological behaviors, the expression of CHI3L1 was reduced in sepsis cell models. As shown in Figure 3A and B, the mRNA and protein expression levels of CHI3L1 in sh-CHI3L1-transfected RAW264.7 cells were decreased. The results of CCK-8 assay revealed that the knockdown of CHI3L1 promoted the viability of LPS-exposed RAW264.7 cells (Fig. 3C). The results of flow cytometry demonstrated that the knockdown of CHI3L1 inhibited the apoptosis of LPS-exposed RAW264.7 cells (Fig. 3D). CHI3L1 knockdown significantly inhibited the levels of inflammatory cytokines (IL-1β, IL-6, and TNF-α) in the LPS-exposed RAW264.7 cells (Fig. 3E-G). The results of western blot analysis revealed that CHI3L1 knockdown inhibited the phosphorylation levels of IκB-α and p65 (Fig. 3H). Then, we further performed an immunofluorescence experiment to demonstrate whether CHI3L1 affects the p65 nuclear translocation. The results showed that LPS treatment promoted P65 nuclear translocation, while CHI3L1 knockout inhibited P65 nuclear translocation (Fig. 3I). These results suggested that the knockdown of CHI3L1 inhibited the activation of the NF-κB pathway. On the whole, these results suggested that the knockdown of CHI3L1 inhibited the inflammation and apoptosis of LPS-exposed RAW264.7 cells by regulating the NF-κB pathway.
Fig. 3
Knockdown of CHI3L1 inhibited sepsis cell apoptosis. A) mRNA expression of CHI3L1 detected by qRT-PCR. B) Cell viability assessed by CCK-8 assay. C) Cell apoptosis analyzed by flow cytometry. Concentrations of inflammatory factors IL-1β (D), IL-6 (E), and TNF-α (F) measured by ELISA. (G) Levels of key enzymes in the NF-κB pathway detected by Western blot. H) Western blot analysis revealed that CHI3L1 knockdown inhibited the phosphorylation levels of IkB-a and p65. I) IF was performed to demonstrate p65 nuclear translocation. **p < 0.01, ##p < 0.01
CHI3L1 knockdown inhibits organ damage in mice with sepsis
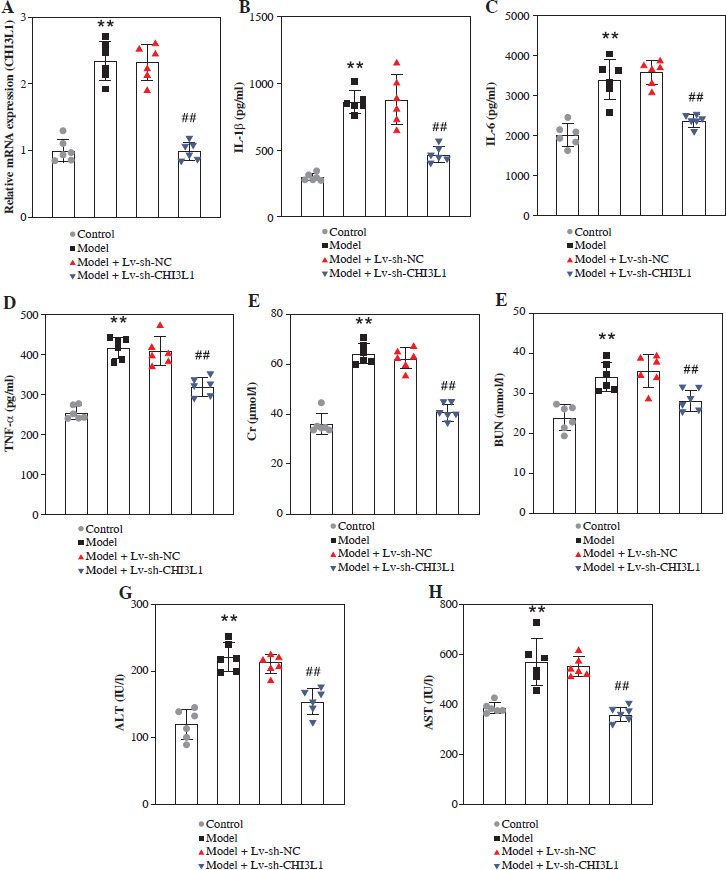
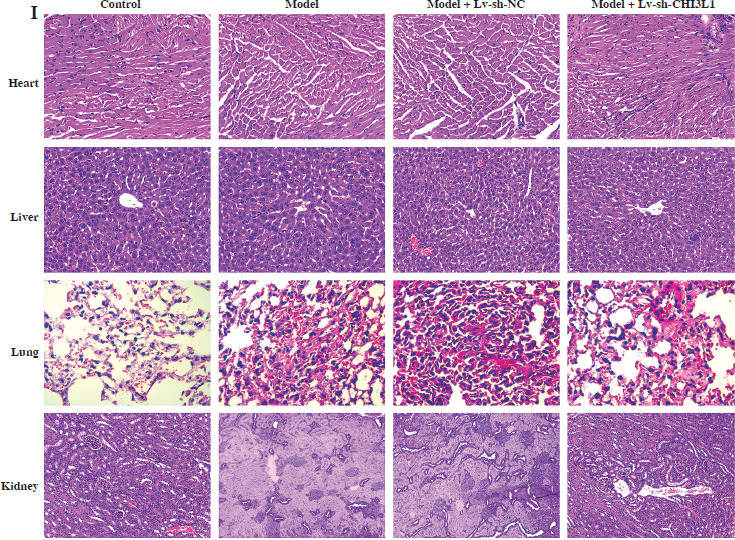
To investigate the effects of CHI3L1 in vivo, a mouse model of sepsis was constructed. First, we detected CHI3L1 levels in all mice of each group. The mRNA level of CHI3L1 was elevated in the serum of sepsis mice, and decreased after Lv-sh-CHI3L injection (Fig. 4A). The results of ELISA revealed that CHI3L1 knockdown significantly reduced the levels of inflammatory cytokines (IL-1β, IL-6, and TNF-α) in the serum of mice in the model + Lv-sh-CHI3L1 group (Fig. 4B-D). Secondly, the contents of Cr, BUN, ALT, and AST in the mice in the model + Lv-sh-CHI3L1 group were significantly lower than the corresponding levels in the other groups (Fig. 4E-H). Finally, H&E staining revealed that as compared with the myocardium in the control and model + Lv-sh-CHI3L1 groups, the myocardium in the model and model + Lv-sh-NC groups exhibited greater disorder in myocardial fiber arrangement, and some of the nuclei in myocardial cells were disrupted. In the liver sections, there was considerable congestion of veins, as well as hepatocyte necrosis in the model and model + Lv-sh-NC groups. In the pulmonary sections, the marked destruction of alveolar structures was detected in the model and model + Lv-sh-NC groups, and in these mice, the effusion in alveoli was markedly more severe than that in the control and model + Lv-sh-CHI3L1 groups. Moreover, tissue infiltration by inflammatory cells was substantially higher in the model and model + Lv-sh-NC groups than in the control and model + Lv-sh-CHI3L1 groups. In the kidney tissue, necrotic glomeruli were considerably more numerous in the model and model + Lv-sh-NC groups than in the control and model + Lv-sh-CHI3L1 groups. In addition, whereas the tubule structure was almost normal in the control and model + Lv-sh-CHI3L1 groups, casts were observed in the tubules in the model and model + Lv-sh-NC groups (Fig. 4I).
Fig. 4
CHI3L1 knockdown inhibited organ damage in sepsis mice. A) CHI3L1 levels in serum of mice detected by qRTPCR. Concentrations of inflammatory factors IL-1β (B), IL-6 (C), and TNF-α (D) measured by ELISA. E-H) Levels of serum creatinine (Cr), blood urea nitrogen (BUN), alanine aminotransferase (ALT), and aspartate transaminase (AST) in mice serum analyzed using an automatic biochemical analyzer. **p < 0.01, ##p < 0.01 I) Analysis of organ damage (heart, liver, lung, kidney) by HE staining in each group of mice
Discussion
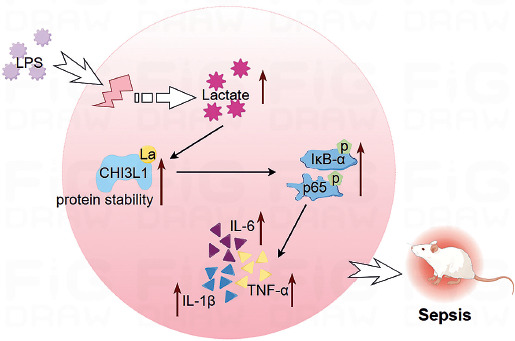
Lactylation is often involved in the development and progression of various diseases. In the present study, LPS-stimulated RAW264.7 cells were used as the sepsis cell model and LPS-injected mice as the animal model. Through in vitro and in vivo experiments, it was confirmed that the lactylation of CHI3L1 promotes the development of sepsis (Fig. 5).
Immune dysfunction is the core mechanism of sepsis. Macrophages are one of the most critical cells that comprise the immune system. The occurrence of sepsis triggers the uncontrolled apoptosis of macrophages, exacerbating inflammatory reactions [25] and promoting the progression of sepsis. CHI3L1, secreted by immune and inflammatory cells, is associated with severe diseases, such as colorectal inflammation, osteoarthritis, and leukemia [11, 26, 27]. Studies have shown that CHI3L1 affects the progression of sepsis [7, 28]. CHI3L1 is closely related to the survival rate of patients with sepsis, skeletal muscle stem cell damage, and the reduced polarization of M2 macrophages [29]. The present study investigated the effects of CHI3L1 on macrophage behaviors in sepsis. CHI3L1 affects disease progression by mediating the apoptosis of cells. Yu et al. [30] found that CHI3L1 promoted the apoptosis of cancer cells, thereby inhibiting the metastasis of cancer cells to the lungs. The study by Higashiyama et al. [31] suggested that CHI3L1 accelerated the progression of liver fibrosis by inhibiting macrophage apoptosis. The apoptotic process of macrophages is accompanied by a large release of inflammatory factors, which contributes to the development of sepsis. Numerous studies have confirmed the effects of CHI3L1 on IL-1β, IL-6, and TNF-α [11, 27, 28, 32]. This is consistent with the findings of the present study. NF-κB is a key signaling pathway that induces cell apoptosis. The study by Yin et al. [33] suggested that the NF-κB pathway mediates macrophage apoptosis. Zhao et al. [34] suggested that CHI3L1 mediates the NF-κB pathway and inhibits the proliferation of glioma cells. The present study found that the knockdown of CHI3L1 mediated the NF-κB pathway and inhibited macrophage apoptosis in models of sepsis.
The N-terminal amino acids of CHI3L1 are composed of 40 kDa of tyrosine, lysine, and leucine [35]. Therefore, it was hypothesized that CHI3L1 could be influenced by PTMs. The study by Yin et al. [36] indicated the presence of m6A methylation modification in CHI3L1. Research by Bhat et al. [37] demonstrated that the acetylation modification of CHI3L1 accelerated the progression of glioma. The present study provides a new insight, demonstrating that the lactylation of CHI3L1 promoted macrophage apoptosis. Zhang et al. observed that, under hypoxic conditions, the incomplete oxidation of glucose in macrophages led to an increase in the production of metabolite lactate. LA generates lactoyl CoA, which provides a lactoyl group to the lysine tail of histones through acyltransferase, resulting in a histone modification called lysine lactylation. This type of PTM affects the function of macrophages by regulating the expression of related genes during proliferative and apoptotic processes [38]. The progression of sepsis is accompanied by the extensive apoptosis of macrophages, which leads to the suppression of immune system function. It causes recurrent bacterial infections and persistent inflammation in patients with sepsis. Yang et al. [19] proposed that histone lactylation modification inhibits the progression of sepsis. In addition, Chu et al. [39] confirmed that the lactylation modification of H3K18la in macrophages mediates the regulation of sepsis development by inflammatory cytokines. The present study identified similar mechanisms. The lactylation modification of CHI3L1 regulates the expression level of inflammatory factors. This process inhibits the progression of sepsis by inhibiting macrophage apoptosis. However, the present study has certain limitations. It did not analyze the specific lactylation modification sites of CHI3L1 or which enzyme regulates the lactylation of CHI3L1. In the future, the authors aim to use bioinformatics analysis and liquid chromatography methods to conduct a more in-depth analysis of these aspects. This will help to comprehensively improve the research findings.
The findings of the present study have several implications for clinical practice, particularly in the context of sepsis management. Understanding the role of CHI3L1 lactylation in promoting the development of sepsis may open new avenues for both diagnosis and treatment. First, given the critical importance of early intervention in improving patient outcomes, identifying patients who are at high risk due to CHI3L1 lactylation levels could enable timely and targeted treatment strategies. Additionally, monitoring changes in CHI3L1 lactylation may provide insight into disease progression and prognosis. Patients with higher levels of lactylated CHI3L1 may have a more severe course of sepsis, indicating the need for more aggressive therapeutic interventions. Furthermore, given the heterogeneity of sepsis presentations, personalized approaches based on the lactylation status of CHI3L1 could lead to more effective treatment protocols. By stratifying patients based on their CHI3L1 lactylation profiles, it may be possible to identify those who would benefit most from specific interventions.
However, there were still some limitations in this study. Whether CHI3L1 has other PTMs (phosphorylation, acetylation, etc.) involved in the progression of sepsis remains to be further explored. In addition, this study used shRNA-mediated CHI3L1 knockdown mice. In the future, we need to further demonstrate our research results in CHI3L1 gene knockout mice.
In conclusion, the present study demonstrated that the LA modification of CHI3L1 promotes macrophage apoptosis by mediating the NF-κB pathway. This process releases a large number of inflammatory cytokines, which promotes the progression of sepsis. These findings provide a basis for the potential role of CHI3L1 as a therapeutic target for sepsis.